
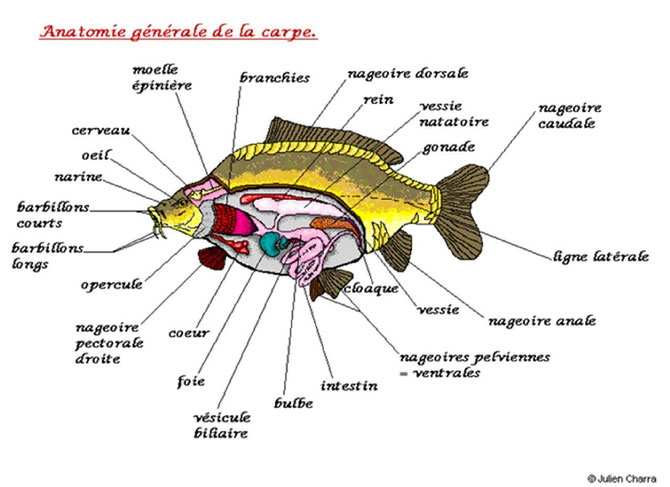
Anatomie générale de la carpe

Appareil respiratoire de la carpe
Les branchies constituent l’échangeur respiratoire principal des poissons permettant d’absorber l’oxygène (O2) dissous dans l’eau et de rejeter le dioxyde de carbone (CO2).
Organisation et structure des branchies :
Les branchies sont formées par des évaginations de la cavité pharyngienne. Elles sont portées par 4 arcs branchiaux chez
les poissons osseux (5 chez les poissons cartilagineux comme les raies et les requins). Sur les arcs branchiaux sont implantées de petites protubérances appelées branchiospines. Les
branchiospines retiennent les particules alimentaires à la manière d’un tamis.
Les branchies sont divisées en filaments branchiaux eux-mêmes divisés en lamelles branchiales. Elles sont richement vascularisées et la barrière épithéliale séparant le sang du milieu extérieur
est très fine (de l’ordre de 1 µm soit 10-6 m).
Elles sont enfermées dans la cavité branchiale, recouverte par une plaque osseuse appelée opercule (chez les poissons osseux uniquement).
Les échanges respiratoires :
Les échanges respiratoires sont régis par 2 principes physiques : la diffusion et la convection.
La diffusion :
La diffusion correspond au transport simple des gaz à travers l’échangeur respiratoire selon des gradients de pression partielle (le gaz passe du milieu où il est le plus concentré vers le milieu où il est le moins concentré). Les flux de gaz transférés par diffusion dépendent de la loi de Fick :
F = - D (S/e) Ç P
F = flux de gaz en mol.s-1
D = coefficient de diffusion en mol.s-1.m-1.Pa-1
S = surface de l’échangeur respiratoire en m2
e = épaisseur de la barrière épithéliale séparant le milieu ambiant du sang en m
Ç P = différence de pression partielle du gaz entre le milieu ambiant et le sang en Pa
La structure de la branchie facilite la diffusion des gaz. D’une part le découpage de la branchie en filaments branchiaux et lamelles branchiales permet d’augmenter la surface d’échange. D’autre part la faible épaisseur de la paroi de la branchie permet un meilleur passage des gaz.
La convection :
La convection correspond au transport des gaz par un fluide.
Il y a un système de convection externe et un système de convection interne.
La convection externe est assurée par les mouvements de la bouche et des opercules qui fonctionnent comme un système de double pompe. La pompe operculaire permet d’aspirer de l’eau par la bouche
alors que la pompe buccale permet d’expulser l’eau par les fentes branchiales.
La convection interne est assurée par la circulation sanguine qui permet de transporter les gaz. Les érythrocytes (globules rouges) des Vertébrés contiennent de l’hémoglobine (un pigment
protéique contenant du Fer) permettant la fixation de l’oxygène. A la différence des érythrocytes des Mammifères, les globules rouges des poissons sont nucléés (ils conservent leur noyau
cellulaire) et ils contiennent plusieurs types d’hémoglobines simultanément (alors que chez les Mammifères il y a différents types d’hémoglobines qui se succèdent au cours du développement :
hémoglobines embryonnaires, fœtales et adultes).
L’affinité de l’hémoglobine pour l’oxygène varie d’une part avec la température et d’autre part avec le pH et la pression partielle de CO2 (effet Bohr). Au niveau d’un tissu en
activité (muscle par exemple) la température et la pression partielle de CO2 sont plus élevées et le pH plus acide. Ces conditions locales particulières facilitent la libération de
l’oxygène. Inversement, au niveau des branchies, la température et la pression partielle de CO2 sont plus faibles et le pH plus basique ce qui facilite la fixation de l’oxygène.
La majeure partie du dioxyde de carbone est transportée par le sang sous forme d’ions bicarbonates (HCO3-). Le reste est transporté sous forme dissoute dans le plasma (mais
contrairement aux Mammifères, Oiseaux et Reptiles il n’y a pas une partie du CO2 liée à l’hémoglobine sous forme de carboxyhémoglobine). La capacité de transport du CO2 par
le sang est inversement proportionnelle à sa teneur en O2 (Effet Haldane).
Effet Bohr et effet Haldane facilitent l’absorption de l’O2 et le rejet du CO2 au niveau des
branchies et les échanges inverses au niveau des tissus.
L’eau circule sur les lamelles branchiales dans la direction opposée à la circulation sanguine. Il s’agit donc d’un système à contre-courant qui facilite les échanges gazeux. L’eau est
progressivement en contact avec du sang de plus en plus désoxygéné alors que le sang est exposé à de l’eau de plus en plus oxygénée ce qui permet de maintenir un gradient de diffusion optimal.
Grâce à ce système à contre-courant les branchies peuvent récupérer 70 à 90 % de l’oxygène dissous dans l’eau.
Besoins en oxygène :
Les besoins en oxygène sont variables d’une espèce de poisson à une autre. Ainsi, la carpe supporte des eaux très pauvres en oxygène (jusqu’à 3ml d’O2 dissous par litre d’eau), alors que la truite commence sérieusement à souffrir lorsque la concentration en oxygène de l’eau passe en dessous de 6ml.L-1. La solubilité de l’oxygène dans l’eau est inversement proportionnelle à la température : dans une eau douce à 0°C, la concentration d’O2 atteint 10 ml. L-1 mais elle chute à 5,6ml.L-1 pour une eau à 30°C. Ceci explique que les truites fréquentent les eaux fraîches de la partie supérieure des rivières et des lacs d’altitude tandis que les carpes se rencontrent plutôt en plaine dans la partie inférieure des rivières, les lacs et les étangs et même de petites mares où l’eau est surchauffée en été. Pourquoi ne rencontre-t-on pas de carpes en altitude ? La carpe peut supporter des températures très basses (jusqu’à 0°C) mais à besoin d’une eau suffisamment chaude pour se reproduire (18-20°c pendant plusieurs jours, conditions qui sont rarement atteintes en altitude).
Autres rôles des branchies :
Les branchies ne sont pas seulement impliquées dans la respiration. Elles interviennent aussi dans l’osmorégulation
(maintien de l’équilibre hydrique et électrolytique de l’organisme).
On appelle osmose la diffusion de l’eau à travers une membrane à perméabilité sélective. Lorsque deux solutions de concentration différente sont séparées par une membrane perméable à l’eau mais
imperméable aux solutés, l’eau a tendance à passer du milieu le moins concentré (hypotonique) vers le milieu le plus concentré (hypertonique) jusqu’à ce que les concentrations de part et d’autre
de la membrane soient identiques (solutions isotoniques).
Chez les poissons d’eau de mer, le milieu intérieur est hypotonique par rapport au milieu extérieur. Ils ont donc tendance à perdre de l’eau par osmose, principalement à travers l’épithélium
branchial. Pour compenser ces pertes d’eau, ils sont obligés de boire de grandes quantités d’eau de mer. L’excès de sel absorbé est éliminé par des cellules spécialisées de l’épithélium
branchial : les ionocytes. Pour limiter les pertes d’eaux, l’urine des poissons de mer est peu abondante et isotonique par rapport à leurs liquides biologiques.
Chez les poissons d’eau douce comme la carpe, le problème est inverse. Le milieu intérieur est hypertonique par rapport au milieu extérieur. L’eau à donc tendance à pénétrer au travers de
l’épithélium branchial. Pour éliminer l’excès d’eau, ils émettent une urine abondante et très diluée (hypotonique par rapport à leurs liquides biologiques). Même si cette urine est diluée, elle
entraîne une perte d’électrolytes importants pour l’organisme. Ces pertes sont compensées par l’absorption active d’ions au niveau des branchies.